�P(gu��n)���҂�
��(j��ng)�����xحCell Metab��(x��)��֬������x�c���Y
����(x��)��ͨ�����д��x��������׃������(x��)����ֳ�����а��Y�Ĺ�ͬ��������(x��)����ֳ��Ҫ֬������ϳ�Ĥ����̖(h��o)���ӡ��������၆��W(xu��)��Ѫ�ܼ����о�����Erin Currie������2013��6����Cell Metabolism�ڿ��ϰl(f��)���}�顰Cellular Fatty Acid Metabolism and Cancer�������£���֬�|(zh��)�ĽǶȌ�(du��)����(x��)�����x�M(j��n)���˾C���������Y(ji��)������֬��������ԁ����ư���(x��)����ֳ������(j��)��ԭ��朽ӣ�http://dx.doi.org/10.1016/j.cmet.2013.05.017��

ǰ��
�M�ܰ��Y����ͺͲ������кܴ�IJ��������(x��)��ͨ�����д��x���������������磬�����Ǵ��xͨ����(hu��)�l(f��)����׃����ʹ���������ėl���£�����(x��)���ǽͽ�һ�ӻ��S��WarburgЧ��(y��ng)����������(x��)����֬���ᣨFA�����x��׃���^���ܵ��P(gu��n)ע��F(xi��n)As��ĩ���Ȼ��͟N朽M�ɣ���Ҫ��ż��(sh��)̼����ʽ���ڣ�������͵ģ�Ҳ�����Dz�͵ġ�����������?j��)?ch��)�桢Ĥ��ֳ����̖(h��o)���Ӻϳ�������ġ����@��҂���(ji��n)Ҫ�����(x��)���еĴ��x�����c(di��n)��FA�ϳɺ̓�(ch��)���;�������⣬�҂�߀�о���һ�Nͨ�^����FA�����ԁ�p������(x��)����ֳ���D(zhu��n)�Ƶ�ģ�͡�
����(x��)���������x�ĸ�׃
���Y�ĸ����ρ��f��һ�N��(x��)�����L(zh��ng)����ֳ���Ɂy�О飬����Ҫ��(x��)���Ļ�����(g��u)��������ᡢ�����|(zh��)��֬�|(zh��)������(x��)������ꐴ��x�����ܵ��ɔ_���@ʹ�������܉�e����ꐴ��x���g�a(ch��n)������@Щ��(g��u)���ā�Դ������(x��)������������Ĵ��x�Ɂy��WarburgЧ��(y��ng)����ʹ����������r��Ҳʹ���ǽͽ�;�����������������ǔzȡ���������ӣ��������ữ�p�٣��Լ�����Įa(ch��n)����
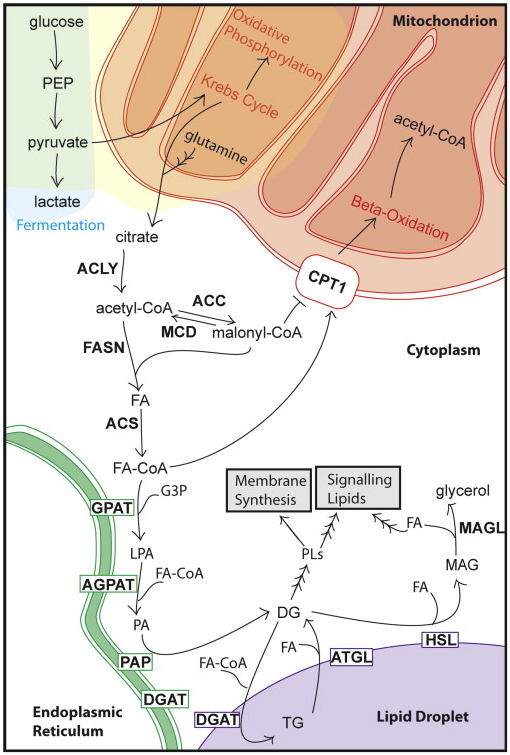
�D1��(x��)��֬������x����
��һ��(g��)��Ҋ�İ��Y���x��׃�ǹȰ��������x���ӡ��ڲ����(d��ng)�:(x��)���У��Ȱ�������һ�N��Ҫ�����������(j��ng)�^���x�����D(zhu��n)�����-ͪ����ᣬ�Ķ��M(j��n)��������ѭ�h(hu��n)���Ȱ����������Ħ�-ͪ�����ͨ�^������ѭ�h(hu��n)�������\(y��n)�D(zhu��n)��(hu��)�a(ch��n)���O����ø��ه�ı�ͪ�ᣬ�����ڙ�����Įa(ch��n)�����Ȱ�����Ҳ�����ڮ�������Ó��ø�͞��^��ø�������£���(j��ng)������ѭ�h(hu��n)�������\(y��n)�D(zhu��n)�a(ch��n)�ə����ᣬȻ��������������a(ch��n)֬����ϳɵ���������
������ֳ��(x��)���е�֬�|(zh��)���xҲ�l(f��)����׃���ڰ���(x��)���У�̼��횏��������a(ch��n)�D(zhu��n)�Ƶ�֬���ᣬ����Ĥ����̖(h��o)���ӵ�����ϳɡ��ּ�(x��)��Ĥ֬�|(zh��)����֬��PLs��������֬��đ�A��PC������֬���Ҵ�����PE�����Լ�����֬�|(zh��)������������֬����ø�w��֬���@Щ֬�|(zh��)�S�ຬ��֬���ᣬ����֬�|(zh��)��Դ�������oøA��֬���ᘋ(g��u)��������Դ����^�ϳɵ�֬���ᡣ�mȻ�����(sh��)������(x��)����ϲ�g��Դ�ԁ�Դ��������(x��)��ƫ�Ï��^�ϳ�֬���ᣬ����(j��ng)�����F(xi��n)����֬����ϳɵ��D(zhu��n)׃�������M(j��n)��������Գأ�֬������Ҫͨ�^֬�������oøA�ϳ�ø�Ĺ��r(ji��)����M(j��n)�С������һ���M(j��n)����Գأ�֬��������c���ͻ����Ǽ��������քe�����������ͣ�TGs����������SEs����Ȼ��(ch��)����֬�Σ�LDs���У�Ҋ�D1�����ڼ�(x��)����(n��i)��֬����������S�����\(y��n)���������Y(ji��)�ϵ�Ĥ�ϡ�����(ch��)�桢׃����̖(h��o)֬�|(zh��)���������ɶ�����̼����������Դ��
�M�ܱ��C����(c��)���ڏ��^�_ʼ��֬����ϳ�;������һЩ���Y����h(hu��n)�������֬�|(zh��)��ʹ֬����zȡ;���ɞ靓�ڵİ��c(di��n)�����磬֬����Y(ji��)�ϵ���4��FABP4������һ�N֬�|(zh��)��H�����c���܇�֬����(x��)�����ѳ�����(x��)���ṩ֬������^�̡����⣬ֻ���ڲ�����֬���ף�һ�N��Դ��֬�|(zh��)Դ������r�����B(y��ng)�r(sh��)��ǰ���ٰ���(x��)���ڴ���FASN��C75����ACLY��SB-204990�����Ƅ�����r���@ʾ�������������͡�CD36��һ�N�V�����_(d��)�Ŀ�Ĥ���ף����ж�N���ܣ�����֬����zȡ���c���ٰ����P(gu��n)�����|(zh��)�M����CD36ˮƽ�����c�[���l(f��)�������ڲ��E���P(gu��n)��ֵ��ע����ǣ���(x��)�����B(y��ng)��(sh��)�(y��n)���w��l�����ܲ�ͬ���w��(n��i)�l����������Դ�Ԕzȡ��ijЩ���Y�п��ܸ�����Ҫ��
ͨ�^����֬���ṩ��(y��ng)�����ư���(x��)����ֳ
����֬���ጦ(du��)����(x��)����ֳ���P(gu��n)��Ҫ����������������ṩһ�N�����ί����ԡ���֬�|(zh��)���x�ĽǶȁ�������֬��������Կ���ͨ�^�N��ʽ��(sh��)�F(xi��n)����1�����֬����ϳɣ���2������֬���������ֽ⣻��3����֬�����D(zhu��n)�Ƶ��惦(ch��)���У���4����ֹ�惦(ch��)���е�֬�����ጷţ��D2�����@Щ���Ʒ�ʽ���ԆΪ�(d��)���ԽM�Ϸ�ʽ��ɣ��Դ˞��ܣ��҂�������cԓģ�����P(gu��n)���C��(j��)��
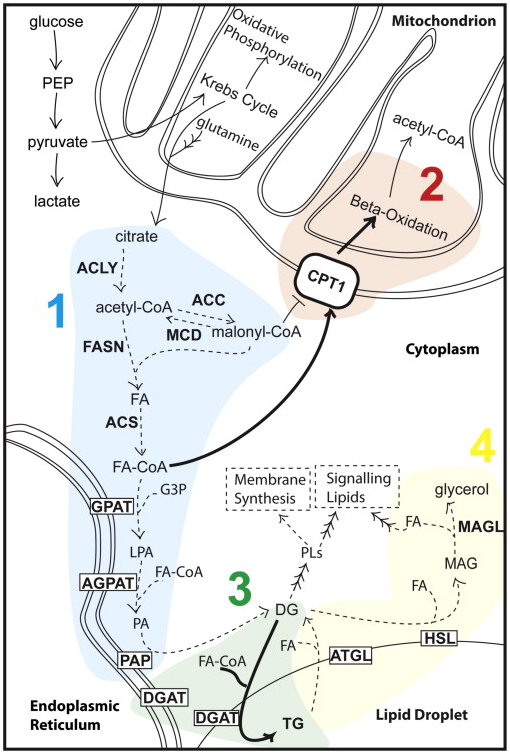
�D2ͨ�^����֬��������ư���(x��)����ֳ��ģ��
���֬����ϳ�
����֬����ˮƽ���(ji��n)�η�������������ĺϳɡ������Ǵ��x�ɱ�ͪ���ͨ�^������ѭ�h(hu��n)���ə����ᣬ�Ķ��M(j��n)��֬������x��Ҋ�D1����̼�ę������D(zhu��n)�����������֬������Ҫ�ׂ�(g��)���E���@Щ���E�漰ATP�������ѽ�ø��ACLY��ACL��ATPCL���������oøA�Ȼ�ø��ACC����֬����ϳ�ø��FASN��FAS���������oøA�ϳ�ø��Ҳ�Q��֬�����oøA�B��ø��ACS��ACSL��FACL�����ڽ���֬��������Ե�ģ���У������@Щø�����ư���(x��)�����L(zh��ng)���@Щ���Ե��R�����x����Ҫ����?y��n)��@Щø�����Ƅ���(du��)�ǰ���(x��)����Ӱ���С��
������ā���(x��)����λ�Q�������Ĵ��x���\(y��n)�������w�еę������(hu��)�M(j��n)��������ѭ�h(hu��n)����(x��)�����|(zh��)�еę������(hu��)����֬����ϳɡ��D(zhu��n)�\(y��n)����CIC���������}�d�w�����{(di��o)��(ji��)�ķ�ʽ�D(zhu��n)�\(y��n)�����ᴩ�^�����w��(n��i)Ĥ���ڼ�(x��)�����|(zh��)��ʹ�á�CICˮƽ�ڸ��N���Y���c�����A(y��)�������P(gu��n)��������������BTA����(du��)�D(zhu��n)�\(y��n)�������ڸ��N�[����ͺͮ��N��ֲС���w��(n��i)�@ʾ���������á�
���֬����ϳɻ���ı��_(d��)
����ֱ�Ӱ�������֬����ϳ�ø�⣬Ҳ����ͨ�^�����D(zhu��n)�ˮƽ������֬����ˮƽ��֬����ϳɵ���Ҫ�D(zhu��n)��{(di��o)�����������{(di��o)��(ji��)Ԫ���Y(ji��)�ϵ���1��SREBP-1����SREBP-1�ЃɷN���ͣ�SREBP-1a�Ǵ����(sh��)���B(y��ng)��(x��)��ϵ�е���Ҫ���ͣ�SREBP-1c�ڸ��K�ʹ����(sh��)�M����ռ����(d��o)��λ��������ˮƽ�£�SREBP-1cͨ�^��(y��ng)�������ACLY��ACC��FAS��SCD-1��GPAT������֬��������ϳ�;������ˣ����ư���(x��)���е�SREBP-1���Խ���֬����ϳɻ���ı��_(d��)��������ֹ����(x��)����ֳ�������Σ�SREBP�D(zhu��n)������ܶ���(g��)��̖(h��o)ͨ·�{(di��o)��(ji��)���������L(zh��ng)������̖(h��o)����(d��o)���KX�������w��LXR����ͨ�^�T��(d��o)SREBP-1c���_(d��)������֬����ϳɡ���ˣ�����ͨ�^����LXR�����p������(x��)����ֳ��Ȼ����LXR�ļ���e��ͨ�^T0901317���������������ٰ����Y(ji��)�c����ǰ���ٰ��а���(x��)������ֳ���@Щ�f��LXR�����{(di��o)��(ji��)FA�ϳ�֮��߀�й��ܡ�
����֬���������ֽ�
���Ͱ���(x��)����֬����ˮƽ�ĵڶ���(g��)�����������併�����ʡ������֬�����ܱ������w�����������x��֬�����oøAͨ�^��A�ؙ����D(zhu��n)��ø1��CPT1���D(zhu��n)����֬������A�İ��{�羀���w��Ĥ�D(zhu��n)�\(y��n)���ھ����w��(n��i)��ÿ�M(j��n)��һ�Φ�-�������ã���(hu��)�ֽ��һ��(g��)��̼Ƭ�Σ������^ԭ���كɂ�(g��)̼ԭ�ӵ�֬����֬���a(ch��n)�������oøA��Ȼ���M(j��n)��������ѭ�h(hu��n)ጷŴ���ATP������֬��������������֬����ˮƽ����Փ�Ͽ��У�����(sh��)�(y��n)��(sh��)��(j��)���y�(y��n)�C��
��֬�����D(zhu��n)�Ƶ��惦(ch��)����
֬���������Ĥ֬�ϳɡ������(ch��)�档������������֬�|(zh��)����TGs����������֬����ă�(ch��)�����Ӻ�����Ĥ��(g��u)������̖(h��o)֬�|(zh��)�Ļ���֬�����(hu��)�p�٣��Ķ����Ƽ�(x��)����ֳ�������(sh��)��(x��)����֬���ღ(ch��)���ڸ��������еİ��|(zh��)֬�Σ�LD���У�ԓ��(x��)��������Ҫ������֬�|(zh��)��(ch��)�档LDs�ڰ���(x��)���е������в�������mȻ��(j��)��(b��o)�����S���(x��)���а��|(zh��)֬�Δ�(sh��)�����ӣ������@�N�۷e���J(r��n)�����²��Եģ������|(zh��)֬�ε��۷e�������ܲ��������?zh��n)ס������������ĬF(xi��n)�ɵ�֬����ؿ������²��Եġ����|(zh��)֬���۷eҲ���ܷ�ӳ��(x��)����(du��)��(y��ng)���ķ���(y��ng)��δ�����о�߀��(y��ng)�м�(x��)�������|(zh��)֬���۷e�ǰl(f��)���ڰ���(x��)����(n��i)߀���܇���(x��)����(n��i)��
��Ҫ�ĸ��������ϳ�;���Q��Kennedy���������;������ʹ�ø���-3-���������D(zhu��n)��ø��GPAT���������������������D(zhu��n)��ø��AGPAT������֬������ˮ��ø��֬����PAP���Ͷ��������������D(zhu��n)�Fø��DGAT����֬�����c����3-����s�ϡ������h(yu��n)��ø��DGAT���⣬����ø�Įa(ch��n)�ﶼ�M(j��n)����֬�ϳɡ���ˣ�GPAT��AGPAT��֬�����ܱ�������������֬�a(ch��n)����������֬���ღ(ch��)���Ŭ���������ڼ���DGAT�����⣬����֬���ღ(ch��)��ĝ�����̎����ֻ����ͬ�r(sh��)����֬����ă�(ch��)����ጷŵ���r�²��܌�(sh��)�F(xi��n)��
��ֹ�惦(ch��)���е�֬�����ጷ�
һ����(ch��)�棬֬����ɱ�ጷŹ��ض�֬��øʹ�á�ͨ�^��ֹ�惦(ch��)����֬���ֽ⣬�����ڰ���(x��)����ֳ�Ļ���֬����ؿ��ܜp�١�֬���ֽ�a(ch��n)����֬����Ҳ��������Ҫ��̖(h��o)֬�|(zh��)��ǰ�w�������(sh��)�P(gu��n)��֬���ֽ��֪�R(sh��)��Դ�ڌ�(du��)֬����(x��)�����о�����������Ó��ø��LD���е�ÿ��(g��)�����������ӿ�ͨ�^֬����������֬��ø��ATGL������������֬��ø��HSL���͆���������֬��ø��MAGL����������ñ���ȫˮ��ጷ����N֬���ᡣ�M���@Щ֬��ø�е�ÿһ�N�������M����Ҳ������Ҫ���ܣ����в��������֬��ø�Ƿ���������(x��)������аl(f��)�]���á�Ŀǰ�������(sh��)�P(gu��n)��֬��ø�Ͱ��Y�Ĕ�(sh��)��(j��)����ᘌ�(du��)����������֬��ø�ġ�
�Y(ji��)Փ
����(x��)����ه֬�������鼚(x��)��������(g��u)��������Ĥ�γɡ�����?j��)?ch��)�����̖(h��o)���ӵĮa(ch��n)�����������(x��)����ֳ��Ҫ֬�����ģ�������_�ģ���ô����(x��)��������֬������x;���д��ڶ���(g��)���c(di��n)�������Ɖİ���(x��)��������ֳ�����@Щ���c(di��n)�ѽ�(j��ng)�挦(du��)��(y��ng)�����Ƅ�����1�������������Ǵ��xһ�ӣ�ᘌ�(du��)֬������x���܌�(du��)����ֳ��(x��)�������x���ԡ����ߣ��Լ�(x��)���خ��ԺͰ����Եķ�ʽ���f��֬������x���Ƅ�Ҳ�ǝ����ί���ʽ��

���Y����ͺ͝��ڵĻ����׃�Ƕ�N���ӵġ�֬�|(zh��)���x���Ǐ�(f��)�s�ģ����S�ͬ�ķ����C(j��)�ƺ��{(di��o)��(ji��)�c(di��n)�����⣬�����(sh��)֬�|(zh��)���xø���ж�N���ͣ��@Щ���Ϳ��c��ͬ��֬�|(zh��)���x�^����ϣ����в�ͬ�ļ�(x��)����λ��M���ֲ�����ˣ��ί���ʽ�ijɹ��c�����ȡ�Q�ڌ�(du��)�ض���Ͱ��Y���ض����x�����C(j��)�Ƶ��˽�̶ȡ�







